ゲノムの塩基配列は交雑、組み換え、共生、感染、水平転移などの様々な要因によって不断に変化しています。そのような変異の中には、エピゲノムの変動を介して植物の多様性や進化に影響を与えるものがあります。私たちのグループでは、シロイヌナズナなどの植物を用いた実験的なアプローチと、コンピュータを用いたゲノム・トランスクリプトーム・エピゲノム・メタゲノムデータの解析の両面から研究を行なっています。また、これらの解析から得られるノウハウを活かした新規解析手法の開発にも取り組み、植物の進化だけでなく、農業にも貢献できる研究を推進しています。
私たちは、主に、以下の3つのテーマで研究を行っています。
- 遺伝子発現やエピジェネティクスの視点による植物の進化研究
- 植物が持つゲノムの進化プロセスを明らかにするためのゲノム系統解析法の開発
- 生態系の系統樹の作成と応用
1. 遺伝子発現やエピジェネティクスの視点から植物の進化を考える
植物の進化 と 転写調節系 の獲得
植物の進化には葉緑体の獲得、水生植物の陸上進出といった興味深いプロセスがたくさん含まれています。このような進化の過程では、ゲノム上で多数の遺伝子の獲得・消失が起こり、さらには転写調節系の獲得・変化のような遺伝子発現レベルの進化も生じます。例えば、葉緑体の起源は植物の祖先生物の細胞内に共生したシアノバクテリアですが、現在の植物の染色体には千以上のシアノバクテリア起源の遺伝子が含まれています。これらは、進化の過程で葉緑体ゲノムから核ゲノムに転移し、そこで新たに転写調節系を獲得したと考えられています。
一方、真核ゲノムには転写される領域と転写されない領域がありますが、この二つを区別する根本的な仕組みには、まだ多くの謎があります。20世紀の生物学では、転写領域はプロモーター配列によって単純に決定されると考えられていましたが、21世紀の今日では、クロマチンの微細構造やDNAの化学修飾などを含む「エピジェネティックな仕組み」が、ゲノムDNAの「読み方」に大きく関わることがわかってきました。
遺伝子の転移によって引き起こされる転写の変動
私たちのグループでは、葉緑体から核に転移した遺伝子の転写調節系の獲得過程に興味を持ち、その仕組みを進化の再現実験とエピジェネティクスの解析手法を用いて研究してきました。具体的には、シロイヌナズナの染色体に、葉緑体遺伝子に見立てた “プロモーターのない遺伝子” を多数導入し、その発現を並列的に解析する方法を開発しました*1*2。そして、遺伝子の発現の強さと染色体上の位置、遺伝子周辺の塩基配列、ヒストン修飾などについて詳細な解析を行いました。その結果、導入した遺伝子のうち、あるものは既存の遺伝子に融合し、別のものはそれまで機能していなかったプロモーター配列を活性化することで転写できるようになることが明らかになってきました*3。また、後者のようなプロモーターの活性化には、ヒストン修飾などのクロマチン構造の変化が関与することもわかってきました*4。
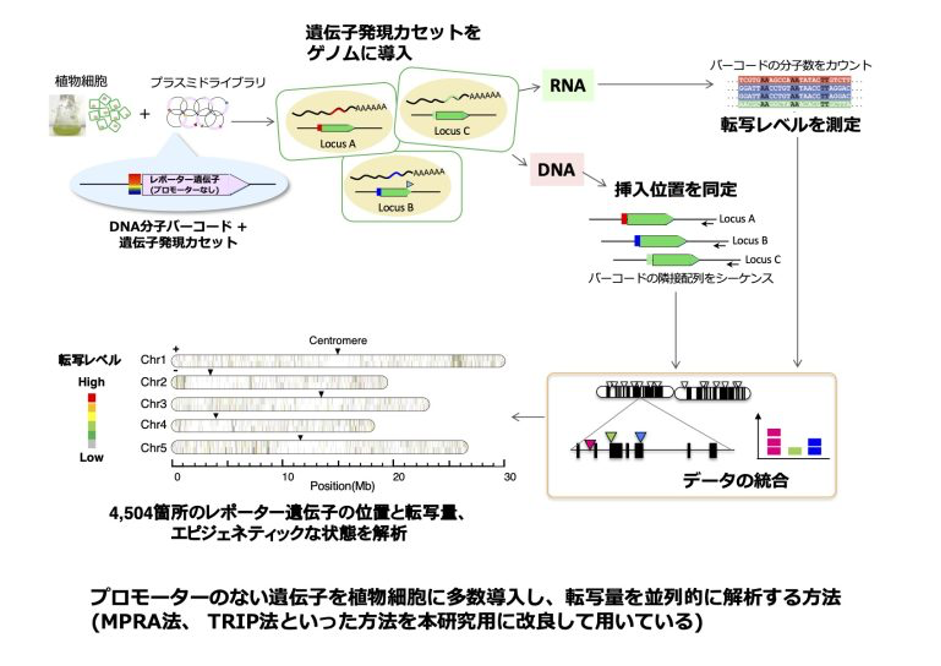
現在私たちは、このような進化の過程で起こる転写変動について、ヒストンの役割やヒストンの分布を変えるメカニズムに着目した研究を行っています。また、このような研究をベースとして、ゲノム編集のような現代の育種・遺伝子操作に応用可能な技術の開発も進めています。
転写に関わるヒストンの変化は何によって引き起こされるのか?
では、プロモーターの活性化につながるヒストンの変化は、何をきっかけに生じたのでしょうか? この疑問について、私たちはDNAの修復機構に着目しました。DNAの修復に関わる因子の変異体の中には、植物の形質転換効率 (遺伝子の導入効率) に作用するものがあることが報告されています。私たちは、シロイヌナズナの細胞内でDNAの切断を誘導できる植物を作成し、DNAの修復時におけるエピジェネティックな変化を解析しました。その結果、DNAの修復に伴い、その周辺領域で変動する修飾ヒストンやヒストンバリアント、RNAポリメラーゼIIによって転写されるRNAがあることがわかってきました*5*6。
2. ゲノム情報から植物の進化プロセスを明らかにする
ゲノム系統樹
植物の進化を調べるにはいくつかの方法があり、化石を調べる、あるいは形状や機能の共通性や地理的な分布を調べることで、進化の歴史を推定することが可能です。これらの方法に対して、遺伝子やタンパク質の配列を比較する方法があります。分子系統樹と呼ばれる進化の歴史を表す図は、一般的に、さまざまな生物間における遺伝子やタンパク質の配列の類似性をもとにして書かれています。
遺伝子の塩基配列は、DNAシーケンサーを用いて調べることができます。2005年くらいからは、次世代シーケンサーと呼ばれる装置が使われるようになり、それまでとは比べ物にならない速度で、さまざまな生物の遺伝子やゲノムの配列が解析されるようになってきました。また、配列データを記録し解析するためのコンピュータの性能や、GenBankやDDBJなどのデータベースに蓄積されるゲノム情報の量も飛躍的に向上し、現在では、多様な生物のゲノム情報を容易に入手し、解析することができるようになりました。こうした時代の流れの中、一つ一つの遺伝子ではなく、ゲノム情報を用いて分子系統樹を書く方法が模索されるようになってきました。例えば、色々な遺伝子のアライメント*7 を連結した仮想巨大アライメントで系統樹を描く方法、多くの系統樹を融合させる方法、ゲノムに含まれる特徴 (GC含量や特定の配列パターン、代謝系遺伝子群…) を比較する方法など、様々な方法が開発されてきました。
*7. 複数の生物種の遺伝子やタンパク質の配列を揃えて並べたもの。変異の位置や共通の配列がわかる。
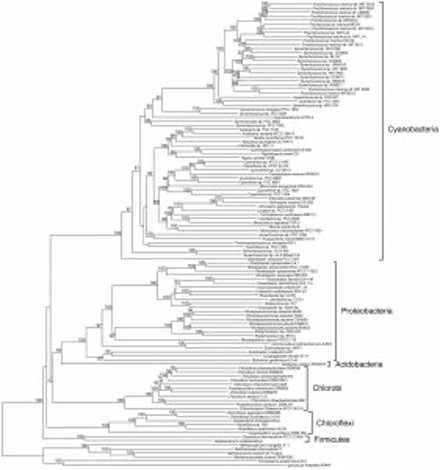
私たちは、遺伝子やタンパク質の類似性を調べるBLASTと呼ばれるツールを利用して、ゲノムに含まれる全遺伝子 (正確には全タンパク質) の配列を比較して分子系統樹を作成する方法を開発しました*8。そして、この方法を用いて光合成生物をはじめとする様々な生物の系統樹を描くことに成功しました*9。このようにして描かれた系統樹には、遺伝子で描かれた系統樹とは異なる進化の系譜も含まれており、今後、新たな発見につながる可能性があります。
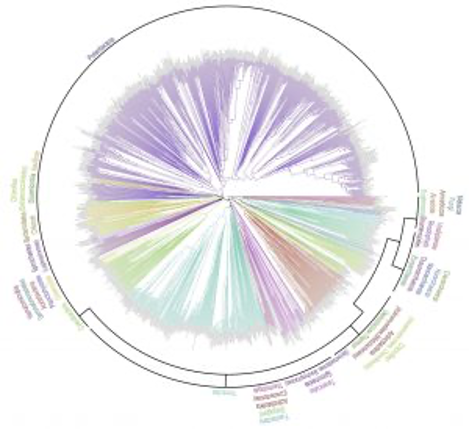
ゲノム系統樹の応用
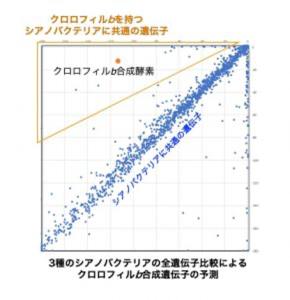
私たちは、ゲノム情報 (全遺伝子配列) を比較し、特定の生物に固有の遺伝子を探索する手法も開発しました。そして、実験的なアプローチが困難な生物種から、光合成色素の合成遺伝子を単離することができました*10*11。また最近、この方法をメタゲノムに応用し。微生物生態系の分析や生態系間の比較解析を行う手法の開発に成功しました(詳しくは次の項目で説明します)。
3. 生物の系統樹から生態系の系統樹へ
メタゲノム系統樹の試み
分子系統樹はいろいろな生物が持つ遺伝子の配列を比較し、似ているものは近縁、似ていないものは遠縁とすることで樹形図を作成する方法であり、遺伝子の視点から生物の進化を見る方法です。これに対してゲノム情報を用いて作る系統樹 (ゲノム系統樹) は、生物の遺伝情報を比較する際に、遺伝子ではなくゲノム全体 (あるいはゲノムに含まれる全て or 多数の遺伝子) を用いる点が従来の分子系統樹と異なっています。
遺伝子の中には進化の過程で塩基配列が変化しやすいものや、*12遠縁の生物のゲノムに種を越えて移動してしまうものがあります。遺伝子の系統樹を描く場合、そのような遺伝子を選んでしまうと実際の生物の進化とはかけ離れた系統樹になってしまうため、系統樹を描く際の遺伝子の選択はとても重要です。一方、ゲノム系統樹は多くの遺伝子を用いるためにそのような問題が起こりにくく、生物(のゲノム)が持つ様々な特徴を余すところなく用いて描いた系統樹であると言えますが、遺伝子の系統樹に比べると開発の歴史が比較的浅いため、様々な方法の開発や理論的な検討が現在も続けられています。
*12. 遺伝子の水平転移や水平移動と呼ばれる現象です。
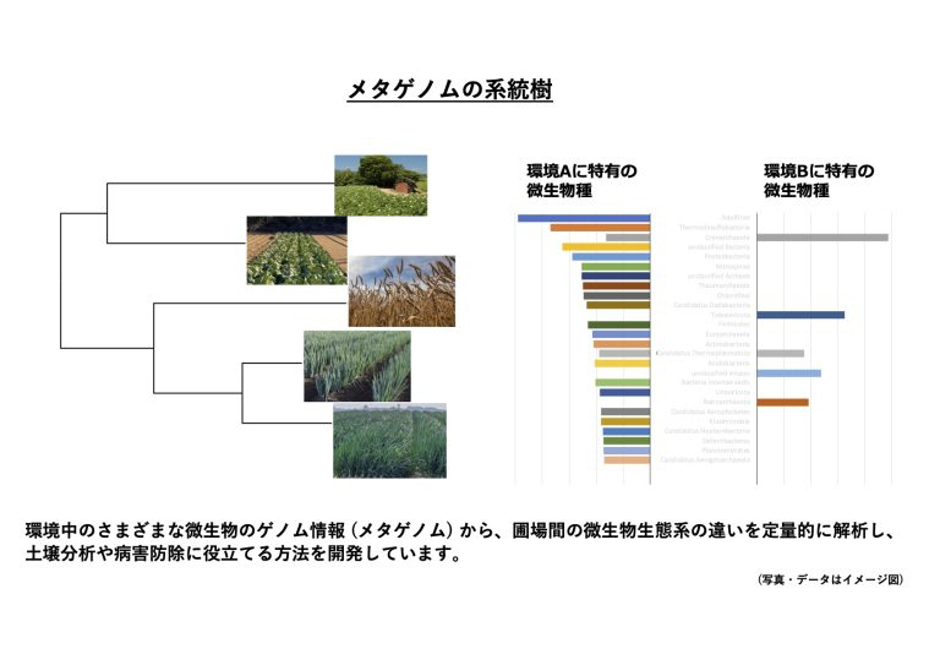
近年、腸内細菌や土壌や水中、さらには大気中や食品といった環境中に含まれる様々な微生物の遺伝子を一つの “ゲノム” としてまとめて解析する研究が盛んに行われるようになり、それらは “メタゲノム” と呼ばれています。例えば、水田の土壌中には嫌気性の細菌が、海面付近には光合成を行う細菌や単細胞藻類が、温泉の泉源付近には高熱に耐性を持つ細菌などが分布していますが、それらを遺伝子やゲノムの視点で調べようという研究分野であり、有用な微生物や遺伝子の発見や、環境の状態のモニタリングなど、様々な応用にもつながっています。
私たちは、これまで生物の進化を表現する方法であったゲノム系統樹を “メタゲノム情報” を用いて作ることで、微生物生態系の変遷(進化)を表現できないだろうか? と思い、メタゲノム系統樹を作成する方法 (MPASS法、 Metagenomic Phylogeny by Average Sequence Similarity) を開発しました*13*14。まだ検討すべきことは多いですが、メタゲノム系統樹を見ることで、微生物群集とそれらを取り巻く環境要因の関係が少しずつ見えてくるようになりました。言い換えると、自然環境中に存在する微生物の種類や量は、温度や水分、微量元素などの量の変動にどのくらい影響されているのか? ということがわかる可能性が出てきたのです。私たちは、この方法の理論や可能性について解析・検討を進めつつ、この方法を農業圃場の土壌のモニタリングなどの応用も含めた研究を行っています。
担当講義
基礎生物学 I(分担,学部1年前期)、生命科学講話(分担,学部1年前期)、遺伝子制御学(学部2年後期)
情報処理基礎演習(農学生命,分担,学部1年前期)、学生実験(学部1〜3年)など